The word "migration" is used in a more familiar sense of the word as meaning a periodical movement of individuals.
Migration, or gene flow, is defined in an evolutionary sense as "the transfer of alleles from the gene pool of one population to the gene pool of another population" (Freeman, Herron 225). Organisms tend to adapt to climate change by changing the geographic distribution of their environmental niche or by altering the specifics of their development and reproduction. This will lead to differences in the compositions of communities and in how they interact with one another (Hoffman). These diverging communities often have different allelic or genotypic frequencies. Gene flow or migration between two populations will help maintain similarities between the species, thus preventing (or slowing down) the process of speciation. In other words, gene flow/migration can help maintain genetic diversity in a population, but it will decrease variation between two populations.
Kirkpatrick and Barton, in their paper "Evolution of a species' range," conclude that gene flow from the center of a species' range can cause maladaptation of organisms that live toward the periphery (implying different environmental conditions) of a species' geographic range. This interference with species adaptation can prevent the species' range from expanding outward. Or even, in some extreme cases, gene flow can produce "demographic sinks" of maladapted individuals in the periphery habitats of their geographic range. Typically, stabilizing selection will act on traits and promote a certain phenotype; the phenotype selected for changes based on the geographic location and environment. Gene flow can actually inhibit this local adaptation due to the re-introduction of alleles into the population. Especially when the environment changes rapidly, this can cause an overall decrease in population size because of a failure to adapt adequately to environmental changes!
Indeed, in subsequent studies by Dawson et al., the researchers conclude that the primary cause of the northern range boundary in the volcano barnacle species Tetraclita rubescens is the "migration load." This migration load arises from the flow of maladapted alleles into the peripheral population. Migration load might be particularly important in species such as barnacles, where reproduction depends on random dispersal of gametes. The researchers infer that environmental change will continue to cause outward (poleward) expansion of geographic ranges, whereas gene flow will continue to set the boundaries on those ranges.
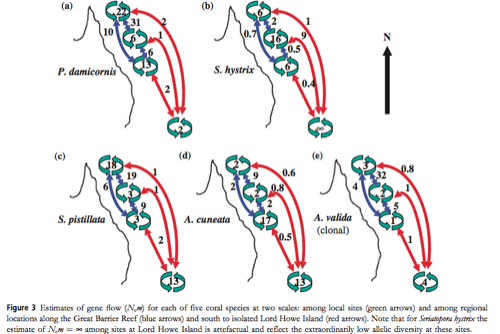
Scientists Ayre and Hughes studied the genotypic diversity and gene flow in several species of coral in order to predict how these species might adapt (or fail to adapt) to climate change (Ecology Letters 2004). They concluded that genetic diversity varies greatly between coral species. The species S. hystrix was found to have low gene flow among local sites and regional sites. This species also had low genetic diversity, and thus the scientists inferred that this species was descended from a very small population initially. Surprisingly, S. hystrix has a very large geographic range. Another species that they studied, S. pistillata, had an equally large geographic range, but it had opposite Fst values (high genotypic diversity). Thus, Ayre and Hughes stated that Fst and Nm values could not accurately predict the geographic range of species. To read more about their research, you can access the paper here: 
The Atlantic cod (Gadus morhuais one such marine animal that has
Back to Introduction
/groups/evolution3000/search/index.rss?sort=modifiedDate&sortDirection=reverse&tag=adaptationlist/groups/evolution3000/search/?sort=modifiedDate&sortDirection=reverse&tag=adaptationAdaptationCustomTagSidebarCustomTagSidebar?sort=modifiedDate&sortDirection=reverse&tag=adaptation0/groups/evolution3000/sidebar/CustomTagSidebarmodifiedDate5CustomTagSidebarreverseadaptationAdaptationcustom/groups/evolution3000/search/index.rss?tag=hotlist/groups/evolution3000/search/?tag=hotWhat’s HotHotListHot!?tag=hot1/groups/evolution3000/sidebar/HotListdheubel5dheubel52012-11-14 16:36:03+00:002012-11-14 16:36:03updated5dheubel5dheubel52012-11-09 16:46:53+00:002012-11-09 16:46:53updated4Added tag - hotdheubel5dheubel52012-11-09 16:46:49+00:002012-11-09 16:46:49addTag3I thought this might be helpful in reading through everything. Let me know if you guys like it!dheubel5dheubel52012-11-09 16:46:24+00:002012-11-09 16:46:24updated2First createddheubel5dheubel52012-11-09 16:31:49+00:002012-11-09 16:31:49created1wiki2012-11-14T16:36:03+00:00groups/evolution3000/wiki/de98eFalseTable of Contents/groups/evolution3000/wiki/de98e/Table_of_Contents.htmldheubel55 updatesTable of Contents
Climate Change
Effects of Climate Change in the Environment
Oxygen Levels
Effects o...Falsedheubel52012-11-14T16:36:03+00:00hot/groups/evolution3000/search/index.rss?sort=modifiedDate&kind=all&sortDirection=reverse&excludePages=wiki/welcomelist/groups/evolution3000/search/?sort=modifiedDate&kind=all&sortDirection=reverse&excludePages=wiki/welcomeRecent ChangesRecentChangesListUpdates?sort=modifiedDate&kind=all&sortDirection=reverse&excludePages=wiki/welcome0/groups/evolution3000/sidebar/RecentChangesListmodifiedDateallRecent ChangesRecentChangesListUpdateswiki/welcomeNo recent changes.reverse5search
Comments